Las pérdidas por esta enfermedad pueden llegar hasta el 80% de la cosecha en años con condiciones climatológicas favorables para su desarrollo
Incidencia de las fuentes de inóculo de 'Monilinia spp.' en la podredumbre parda del melocotonero en poscosecha
M. Villarino; I. Gell; P. Melgarejo; A. De Cal (Departamento de Protección Vegetal, INIA) J. Usall; N.Lamarca (Unidad de Postcosecha del CeRTA, Centro UdL-IRTA)J. Segarra (Departamento de Producción Vegetal y Ciencias Forestales, Universitat de Lleida)02/08/2011
La podredumbre parda del melocotonero constituye una de las enfermedades más importantes de este cultivo y de otros frutales de hueso (por ejemplo: albaricoquero, almendro, cerezo, ciruelo, ) y pepita (manzano, membrillero, peral); y se encuentra presente en todas las zonas donde se cultiva melocotonero en España (De Cal y Melgarejo, 2000).
Introducción
Las pérdidas más importantes se dan en los frutos, pudiendo llegar hasta el 80% de la cosecha en años con condiciones climatológicas favorables para el desarrollo de la enfermedad, sobre todo en huertos de cultivares tardíos (Gell y col., 2008). Los daños en flores son poco importantes en España (Figura 1). Lo mismo ocurre con los daños en brotes y madera (Figura 2), aunque en ocasiones se produce un debilitamiento importante del árbol que conduce a una menor producción y, en algunos casos, a la muerte del mismo.
Figura 1: Marchitez de flores y exudado.
Figura 2: Marchitez de brotes.
El fruto puede ser infectado en cualquier momento de su desarrollo, pero normalmente la gravedad de la enfermedad no es importante hasta que el fruto comienza a madurar (Figura 3), que en el caso de las variedades tardías se inicia seis semanas antes de la cosecha (Villarino y col., 2011). En frutos inmaduros las infecciones pueden permanecer latentes sin mostrar síntomas hasta la madurez.
Figura 3: Podredumbre de fruto en el árbol.
Los primeros síntomas en frutos maduros consisten en pequeñas manchas marrones, que rápidamente muestran podredumbre (Figura 3). Sobre estas zonas se desarrollan pústulas que contienen conidias (Figura 4). Los frutos maduros se infectan directamente de otros frutos, o de conidias dispersadas por el aire que proceden de otros tejidos infectados. Normalmente el fruto infectado permanece sobre el árbol (en los brotes enfermos no se produce la abscisión entre el pedúnculo y el fruto) perdiendo su contenido en agua y dando lugar a momias características de la enfermedad.
Figura 4: Conidias de Monilinia spp. (x100).
Hasta hace unos años únicamente 'Monilinia laxa' (Aderhold & Ruhland) Honey y 'M. fructigena' Honey ex. Whetzel, eran los hongos causantes de la necrosis y podredumbre de los frutales de hueso y de pepita en España (Figura 5). Desde el año 2001, una tercera especie, 'M. fructicola' (Winter) Honey, fue detectada por primera vez en Francia y posteriormente en la República Checa, Italia, España, Eslovenia, Suiza, Alemania, Hungría, Polonia y Rumania (EFSA 2011). Esta especie se encuentra incluida en la lista A2 de organismos de cuarentena en la Unión Europea [i.e., organismos localizados en la región EPPO (European Plant Protection Organization), pero mantenida bajo control oficial] (OEPP/EPPO, 2007).
Figura 5: Especies de Monilinia que causan podredumbre en fruta de hueso.
Materiales y métodos
Se ha estudiado la incidencia del inóculo primario y secundario de las tres especies de 'Monilinia' sobre el desarrollo de la podredumbre de los melocotones después de la cosecha.
Para ello se han realizado quince ensayos durante seis años (2003 a 2008), en huertos de melocotoneros y nectarinas de la zona del valle del Ebro, con los siguientes cultivares: nectarina Autumm Free y Caldesi 20-20, y melocotón Rojo de Albesa, Plácido, O´Henry y Catherine. Se recogieron restos y estructuras vegetales antes de la floración y flores y frutos en ocho fechas diferentes del periodo de cultivo para estudiar la dinámica del inóculo del patógeno en cada una de ellas. Se determinaron las estructuras de supervivencia invernal del patógeno y durante el cultivo se estimaron las infecciones latentes (Aulic), las conidias del patógeno en aire (AUncaC) (Figura 6) y sobre la superficie de la fruta (AUncPC), la podredumbre del melocotonero el día de la cosecha y después de 7 días de postcosecha.
Figura 6: Captador de esporas funcionando en un huerto de melocotones en el Valle del Ebro.
Resultados y discusión
Las tres especies de 'Monilinia' invernan mediante micelio en los frutos momificados que se encuentran en el árbol (Tabla 1) o sobre el suelo (Tabla 2), o en los brotes y ramas infectadas (Gell y col., 2009; Villarino y col. 2010). Dichos materiales sirven de fuente de inóculo primario y cuando las condiciones climáticas son adecuadas, producen conidias que infectan de nuevo flores, brotes y yemas (Byrde y Willets, 1977; Ogawa y col., 1985). Se ha demostrado que existe una correlación positiva entre el número de momias en los árboles y la incidencia de podredumbre de frutos en postcosecha (Tabla 3). El número de momias en los árboles al inicio de la primavera llega a explicar el 65% de la podredumbre causadas por 'Monilinia spp.' en postcosecha (Villarino y col., 2010). Así una momia infectada con 'Monilinia spp.' que haya pasado el invierno sobre cada uno de los árboles de un huerto puede llegar a causar la podredumbre de toda la producción.
Tabla 1: Cuantificación del inóculo primario de Monilinia spp (MO) en el árbol de 8 huertos comerciales del Valle del Ebro. Los datos son la media de las repeticiones de los 30 árboles marcados en cada huerto. Las medias con la misma letra no son significativamente diferentes por el test de Student Newman (SME=cuadrado medio del error) o Kruskal-Wallis (P=0,05). NS= no significativo. (-) No se realizó muestreo.
Tabla 2: Cuantificación del inóculo primario/m2 de Monilinia spp (MO) en el suelo de 8 huertos del Valle del Ebro. Los datos son la media de las 20 repeticiones (20 puntos en el suelo para la toma de muestras) en cada huerto. Las medias con la misma letra no son significativas por el test de Kruskal-Wallis (P=0,05). PB= ramas de poda. NS= no significativo. (-) No se realizó muestreo.
Se ha demostrado también que existe una correlación positiva entre el número de conidias del patógeno por m3 de aire a lo largo del cultivo y el número de momias y ramas infectadas con el patógeno y presentes en el suelo de los huertos al inicio de la primavera (Tabla 3) (Villarino y col., 2010).
Por otra parte, 'M. fructicola' produce otro tipo de inóculo primario, ascosporas en apotecios formados en los frutos momificados que permanecen sobre el suelo de la plantación en el momento de la floración. Sin embargo, por el momento, el principal inóculo primario en los huertos de frutales españoles lo constituyen el micelio y las conidias presentes en las momias que se encuentran en los árboles afectados. No se han detectado apotecios ni ascosporas en las momias del suelo de los huertos del Valle del Ebro (Villarino y col., 2010). Tampoco se ha detectado el patógeno en huesos, restos de hojas y malas hierbas del suelo de los huertos.
Tabla 3: Coeficientes de correlación entre las fuentes de inóculo primario y la podredumbre en postcosecha, infecciones latentes, densidad de conidias de Monilinia spp. (MO) en el aire y sobre la superficie del fruto. Los datos son la media de 10 repeticiones en cada huerto. El dato de porcentaje de podredumbre postcosecha se transformó a arcoseno antes de llevar a cabo los análisis de correlación. Los valores son coeficientes de correlación con sus respectivos valores de P entre paréntesis.
'Monilinia spp.' es un patógeno policíclico en el que se producen numerosos ciclos secundarios a lo largo del ciclo anual de crecimiento del melocotonero (Byrde y Willets, 1977). Las conidias se dispersan por el aire, el agua o los insectos (Luo y col., 2001). Las infecciones secundarias (bien activas o bien latentes) de 'Monilinia spp.' proceden del inóculo producido tras la multiplicación del hongo en las infecciones primarias (Byrde y Willetts, 1977) y aparecen en el Valle del Ebro en todas las fases del desarrollo de la enfermedad: flor, fruto verde y fruto maduro.
Tras la infección de las flores, y particularmente en condiciones de humedad relativa elevada o humectación de los tejidos, el micelio de 'Monilinia spp.' en el tejido vegetal produce hifas cortas que se reúnen, ejercen presión sobre la epidermis, y salen al exterior formando numerosos esporodoquios conidiales sobre los restos florales, desde donde se liberaran nuevas conidias para ciclos secundarios de infección. Al mismo tiempo, el micelio avanza rápidamente hacia el fruto recién formado y hacia el brote. Si la enfermedad progresa en los brotes se forman chancros que pueden llegar a ocupar todo el perímetro de aquéllos y originar la muerte del tejido superior por el estrangulamiento producido.
La invasión de los frutos tiene lugar a través de heridas o aperturas naturales por el micelio producido con la germinación de las conidias, tras lo cual los tejidos son colonizados con rapidez. El patógeno va produciendo pústulas conídicas en las zonas maceradas conforme coloniza el fruto. La dispersión secundaria se debe a una nueva producción de conidias que puede tener lugar 5-7 días después de la infección. En resumen, constituyen fuentes de inóculo secundario las flores, brotes y madera infectada, los frutos de aclareo, así como los frutos enfermos que pueden permanecer o no en los árboles.
Un fruto infectado puede pudrirse en pocos días y, o bien cae al suelo (Figura 7), o permanece unido al árbol. Los frutos que caen generalmente se descomponen. Sin embargo, los que quedan en el árbol, se secan, se arrugan y se convierten en las momias características de la enfermedad. Una vez que el fruto se ha momificado, puede caer al suelo y en muchos casos sobrevivir al ataque de los microorganismos descomponedores, pudiendo persistir el patógeno allí durante casi 1 año, incluso en periodos secos y muy calurosos.
Figura 7: Podredumbre de frutos en el suelo.
La infección de los frutos se produce también después de la cosecha, durante su transporte y almacenaje (Landgraf y Zehr, 1982). Existe una correlación positiva entre la incidencia de la podredumbre del melocotón en el momento de la cosecha y la incidencia de la enfermedad en postcosecha. La incidencia de la podredumbre en el momento de la cosecha puede llegar a explicar el 72% de la incidencia de la podredumbre en postcosecha, según muestra la siguiente ecuación de regresión (R2=0,72; P=0,01) donde debajo de cada uno de los coeficientes de regresión estimados se muestran entre paréntesis sus errores estándard y los porcentajes de incidencia de la podedumbre han sido arcoseno transformados para conseguir la homogeneidad de las varianzas.
Arcsin (% incidencia postcosecha) = 24,6 + 4,05 Arcsin (% incidencia el día de la cosecha)
(4,92) (1,13)
Cuando las condiciones climáticas son desfavorables, las infecciones en frutos pueden permanecer latentes hasta la madurez de éstos, manteniendo al patógeno desde la primavera temprana hasta el momento óptimo de desarrollo de la enfermedad (Byrde y Willetts, 1977). Se ha demostrado que existe una correlación positiva entre el número de infecciones latentes de 'Monilinia spp.' y la incidencia de Podredumbre en postcosecha (Emery y col., 2000; Gell y col., 2008; Luo y col., 2001). Las infecciones latentes llegan a explicar el 54,6% de la incidencia de la enfermedad causada por 'Monilinia spp.' en el Valle del Ebro (Gell y col., 2008).
La presencia de conidias sobre la superficie de los frutos durante el ciclo de cultivo tiene una correlación positiva (r=0,69) con la incidencia de las infecciones latentes (Gell y col., 2009). La densidad de conidias de 'M. laxa' y 'M. fructigena' en los huertos de melocotoneros en España se ve afectada por factores climáticos como la temperatura, la radiación solar, la precipitación y la velocidad del viento (Gell y col., 2009).
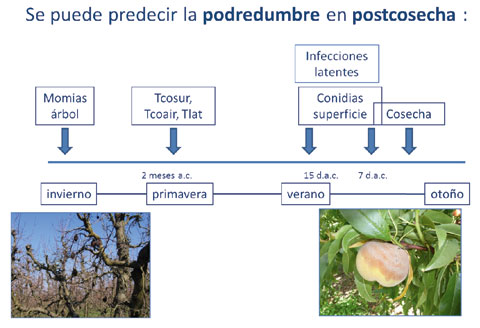
La incidencia de la podredumbre en postcosecha puede predecirse desde el momento en que aparece la primera infección latente, y se observan las primeras conidias del patógeno sobre la superficie del fruto y en el aire. Las fechas en las que aparecen la primera infección latente (Tlat), la primera conidia del patógeno sobre la superficie del fruto (Tcosur) y en el aire (Tcoair) llegan a explicar el 99% de la incidencia de la enfermedad causada por 'Monilinia spp.' en postcosecha.
La cantidad de conidias aéreas presentes en los huertos y sobre la superficie de los frutos varía en el Valle del Ebro, según el año de muestreo y también en función del estado fenológico del cultivo, siendo máxima en la época de cosecha. La densidad del patógeno sobre la superficie de los frutos durante las dos últimas semanas antes de la cosecha, explican el 83% de la podredumbre de estos frutos en postcosecha.
Las condiciones climáticas son críticas para la infección de los frutos, siendo temperatura (T) y periodo de humectación (W), los factores climáticos que más influyen sobre la penetración e infección de los frutos por 'Monilinia spp.' (Gell y col., 2008). En el caso de infección de frutos inmaduros, que generalmente da lugar a infecciones que permanecen latentes, estos dos factores climáticos explican más del 90% de la variación de su incidencia (Gell y col., 2008), teniendo más influencia la duración del período de humectación que la temperatura (Figura 8). En el valle del Ebro, el desarrollo de las infecciones latentes requiere temperaturas suaves y periodos de humectación superiores a 7 h (Figura 8), valores que pueden ser alcanzados durante la primavera tardía y el final del verano. En el caso de podredumbre de frutos en recolección, la incidencia de la enfermedad aumenta con la temperatura y la duración del periodo de humectación. La temperatura óptima para la infección de melocotones por 'M. fructicola' durante la recolección es de 22,5 a 25 °C, con las que más del 79% de los frutos se infectan con periodos de humectación de al menos 12 h y temperaturas inferiores a 27,5 °C.
Figura 8: Superficie obtenida del modelo de regresión múltiple que permite predecir la incidencia de las infecciones latentes en función de la temperatura (T) y del periodo de humectación (W) (Gell y col., 2008).
Conclusiones
El número de frutos momificados presentes en los árboles, las infecciones latentes, la densidad de inóculo del patógeno en el aire y sobre la superficie de los frutos, el estado fenológico del fruto, los valores de temperatura y periodo de humectación, son críticos para la elección de una estrategia de control de la podredumbre del melocotonero.
Referencias bibliográficas
- Byrde, R. J., y Willetts, H. J. 1977. The Brown Rot Fungi of Fruit - Their Biology and Control. Pergamon Press, Oxford.
- De Cal, A. y Melgarejo, P. 2000. Momificado de los frutales de hueso (Monilinia spp.). En: Enfermedades de los frutales de pepita y de hueso. E. Montesinos, P. Melgarejo, M.A. Cambra y J. Pinochet (eds). Ed. Mundi-Prensa. Pp 66-67.
- EFSA Panel on Plant Health (PLH); Pest risk assessment of Monilinia fructicola for the EU territory and identification and evaluation of risk management options. EFSA Journal 2011;9(4):2119. [155 pp.] doi:10.2903/j.efsa.2011.2119. Available online: www.efsa.europa.eu/efsajournal
- Emery, K. M., Michailides, T. J. y Scherm, H. (2000). Incidence of latent infection of immature peach fruit by Monilinia fructicola and relationship to brown rot in Georgia. Plant Disease 84, 853-857.
- EPPO 2007. List of A2 pests regulated as quarantine pests in the EPPO region. OEPP/EPPO from http://www.eppo.org/QUARANTINE/listA2.htm.
- Gell, I., De Cal, A., Torres, R., Usall, J., y Melgarejo, P. 2008. Relationship between the incidence of latent infections caused by Monilinia spp. and the incidence of brown rot of peach fruit: Factors affecting latent infection. European Journal of Plant Pathology 121:487-498.
- Gell, I., De Cal, A., Torres, R., Usall, J., y Melgarejo, P. 2009. Conidial density of Monilinia spp. on peach fruit surfaces in relation to the incidences of latent infections and brown rot. European Journal of Plant Pathology 123:415-424.
- Landgraf, F. A., y Zehr, E. I. 1982. Inoculum sources for Monilinia fructicola in South Carolina peach orchards. Phytopathology 72:185-190.
- Luo, Y. y Michailides, T. J. 2001. Factors affecting latent infection of prune fruit by Monilinia fructicola. Phytopathology 91: 864-872.
- Ogawa, J. M., Zehr, E. I., y Biggs, A. R. 1995. Brown Rot. Pages 7-10 in: Compendium of Stone Fruit Disease. M. Ogawa, E. I. Zehr, G. W. Bird, D. F. Ritchie, K. Uriu, and J. K. Uyemoto, eds. American Phytopathological Society, St. Paul, MN.
- Villarino, M., Melgarejo, P., Usall, J., Segarra, J., y De Cal, A. 2010. Primary inoculum sources of Monilinia spp. in Spanish peach orchards and their relative importance in Brown rot. Plant Disease 94: 1048-1054.
- Villarino, M., Sandin-España, P., Melgarejo, P., y De Cal, A. 2011. High Chlorogenic and Neochlorogenic Acid Levels in Immature Peach Reduce Monilinia laxa Infection by Interfering with Fungal Melanin Biosynthesis. Journal of Agricultural and Food Chemistry 59 (7), 32053213.



")


 en el árbol de 8 huertos comerciales del Valle del Ebro...")
 en el suelo de 8 huertos del Valle del Ebro...")