Brown rot of the peach tree is one of the most important diseases of this crop and other fruits of bone (for example: apricot tree, almond, cherry, Plum,) and pepita (manzano, membrillero, peral); and it is present in all areas where peach is cultivated in Spain (of Cal y Melgarejo, 2000).
Introduction
The most important losses occur in the fruit, being able to get up to 80% of the harvest in years with climatological conditions favourable to the development of the disease, especially in orchards of cultivars delayed (Gell and col., 2008). Damage to flowers are unimportant in Spain (Figure 1). The same applies to damage shoots and wood (Figure 2), although on occasions produced a major weakening of the tree that leads to lower production and, in some cases, the death of the same.
Figure 1: Wilt of flowers and exudate.
Figure 2: Wilt of outbreaks.
The fruit can be infected at any stage of their development, but normally the severity of the disease is not important until the fruit begins to ripen (Figure 3), in the case of the late varieties begins six weeks before harvest (Villarino and col., 2011). In immature fruit infections may remain latent without showing symptoms until adulthood.
Figure 3: Rot of fruit on the tree.
The first symptoms in ripe fruits consist of small brown spots, showing quickly rot (Figure 3). On these areas develop pustules containing conidia (Figure 4). The ripe fruits are infected directly from other fruits, or conidia dispersed through the air from other infected tissues. Normally the infected fruit remains on the tree (in sick outbreaks there is between the stem and fruit abscission) losing its content in water and giving rise to characteristic of the disease mummies.
Figure 4: Conidias of Monilinia spp.. (x 100).
Until a few years ago only 'Monilinia laxa' (Aderhold & Ruhland) Honey and 'M. fructigena' former Honey. Whetzel, were the causative fungi of necrosis and rotting fruit Nugget in Spain (Figure 5) and bone. Since 2001, a third species, 'M. fructicola' (Winter) Honey, was detected for the first time in France and later Czech Republic, Italy, Spain, Slovenia, Switzerland, Germany, Hungary, Poland and Romania (EFSA 2011). This species is included in list A2 quarantine organisms in the European Union [i.e., agencies located in the region, EPPO ('European Plant Protection Organization'), but kept under official control] (OEPP/EPPO, 2007).
Figure 5: Monilinia species that cause rot in stone fruit.
Materials and methods
The incidence of the inoculum primary and secondary of the three species of 'Monilinia' on the development of rot of peaches after harvest has been studied.
This Fifteen trials have been conducted for six years (2003 to 2008), with orchards of peach and nectarines in the area of the Ebro Valley, with the following cultivars: Nectarine 'Autumm Free' and ' Caldesi 20-20', and 'Red of Albesa', 'Placido' peach 'O´Henry', 'Catherine'. Plant structures before flowering and flowers and fruits in eight dates of the period of cultivation and remains were collected to study the dynamics of the inoculum of the pathogen in each one of them. The structures of winter survival of the pathogen were determined and during cultivation estimated latent infections (Aulic) the conidia of the pathogen in air (AUncaC) (Figure 6) and on the surface of the fruit (AUncPC), the rot of the peach tree day of harvest, and after 7 days of post-harvest.
Figure 6: Spores Captador working in a garden of peaches in the Ebro Valley.
Results and discussion
The three species of 'monilinia' wintering through mycelium in mummified fruit on the tree (see table 1) or on the ground (table 2), or infected branches and buds (Gell and col., 2009;) (Villarino and col. 2010). These materials serve as a source of primary inoculum and when weather conditions are suitable, produce conidia that infect new flowers, buds and shoots (Byrde and Willetts, 1977;) (Ogawa et al., 1985). It has shown that there is a positive correlation between the number of mummies in the trees and the incidence of rotting fruit in post-harvest (table 3). The number of mummies in the trees at the start of the spring arrives to explain 65 per cent of the rot caused by ' Monilinia spp. ' on post-harvest (Villarino et al., 2010). So an infected Mummy with ' Monilinia spp. ' has spent the winter on each of the trees in an orchard can cause rot the entire production.
Table 1: quantification of primary inoculum of Monilinia spp. (MO) in tree 8 commercial orchards in the Valley of the Ebro. The data are the average of the repetitions of 30 trees marked in each garden. Measures with the same letter are not significantly different for the test of Student Newman (EMS = square of the error) or Kruskal-Wallis (P = 0, 05). NS = not significant. (-) No sampling was performed.
Table 2: quantification of the inoculum primary/m2 of Monilinia spp. (MO) on the floor of 8 gardens from the Ebro Valley. The data are the average of 20 repetitions (20 points in the soil for sampling) in each garden. Measures with the same letter are not significant for the Kruskal-Wallis test (P = 0, 05). BP = pruning branches. NS = not significant. (-) Sampling was not conducted.
It has also been shown that there is a positive correlation between the number of conidia of the pathogen by m3 of air along the crop and the number of Mummies and infected with the pathogen and branches present in the soil of gardens at the beginning of the spring (table 3) (Villarino et al., 2010).
On the other hand, 'M. fructicola' produces another type of primary inoculum, ascospores in apothecia trained in mummified fruit which remain on the floor of the plantation at the time of flowering. However, for the time being, the main primary inoculum in the Spanish fruit orchards are the mycelium and conidia present in mummies found in affected trees. Apothecia or ascospores in the mummies of the floor of the gardens from the Ebro Valley (Villarino et al., 2010) has not been detected. Neither the pathogen in bones, remains of leaves and weeds from the soil of gardens has been detected.
Table 3: correlation coefficients between the sources of primary inoculum and the rot in post-harvest, latent infections, density of conidia of Monilinia spp.. (MO) in the air and on the surface of the fruit. The data are the average of 10 repetitions in each garden. The data on percentage of post-harvest rot was transformed to arcoseno before carrying out the analysis of correlation. The values are coefficients of correlation with their respective values of P in parentheses.
' Monilinia spp. ' is a polycyclic pathogen producing numerous secondary cycles during the annual cycle of growth of the peach tree (Byrde and Willetts, 1977). The conidia are dispersed through air, water or insects (Luo et al., 2001). Secondary infections (either active or rather latent) of ' Monilinia spp. ' come from the inoculum produced after the proliferation of the fungus in primary infection (Byrde and Willetts, 1977) and appear in the Ebro Valley in all phases of the development of the disease: flower, green fruit and ripe fruit.
After infection of the flowers, and in particular in conditions of high humidity or moisture of tissues, the mycelium of ' Monilinia spp. ' in the tissue plant produces short hyphae meet, put pressure on the skin which leave abroad forming numerous conidiales esporodoquios on floral remnants, from where freed conidia news for secondary cycles of infection. At the same time, the mycelium is moving rapidly towards the newly formed fruit and the outbreak. If the disease progresses in the buds form along that may occupy the entire perimeter of those and thus result in the death of the tissue produced strangulation of more than.
The invasion of the fruit takes place through wounds or natural openings by the mycelia produced with the conidia germination, whereupon the tissues are colonized with speed. The pathogen is producing pustules conídicas marinated areas as it colonises the fruit. Secondary scattering due to a new production of conidia that may take place 5-7 days after infection. In summary, are sources of secondary inoculum flowers, buds and infected wood, the fruits of clearing, as well as the diseased fruits that can stay or not in the trees.
Infected fruit can rot in a few days, and either falls to the floor (Figure 7), or remains attached to the tree. The fruits that fall generally break down. However, remaining on the tree, dries will wrinkle and become features of the disease mummies. Once the fruit has been mummified, it can fall to the ground and in many cases survive the attack of microorganisms decomposers, and can persist the pathogen there for almost 1 year, even in periods of dry and very hot.
Figure 7: Rot of fruit on the ground.
Fruit infection also occurs after the harvest, during transport and storage (Landgraf and Zehr, 1982). There is a positive correlation between the incidence of the rot of the peach in the time of the harvest and post-harvest disease incidence. The incidence of the rot at the time of the harvest may explain 72% of the incidence of post-harvest rot, according to the following regression equation (R2= 0.72;) P = 0, 01) where their standard errors are shown parentheses below for each of the estimated regression coefficients and rates of incidence of the podedumbre have been arcoseno processed to achieve the homogeneity of variances.
When weather conditions are unfavourable, infections in fruits can remain latent until maturity they maintain the pathogen from early spring until the optimum development of the disease (Byrde and Willetts, 1977). It has been shown that there is a positive correlation between the number of latent infections of ' Monilinia spp. ' and the incidence of rot in post-harvest (Emery and col., 2000;) Gell and col., 2008; (Luo et al., 2001). Latent infections reach explain 54.6% of the incidence of the disease caused by ' Monilinia spp. ' in the Valley of the Ebro (Gell et al., 2008).
The presence of conidia on the surface of the fruit during the crop cycle has a positive correlation (r = 0, 69) with the incidence of latent infections (Gell et al., 2009). The density of conidia of 'M. lax' and 'M. fructigena' in the orchards of peach trees in Spain affected by climatic factors such as temperature, solar radiation, precipitation, and the speed of the wind (Gell et al., 2009).
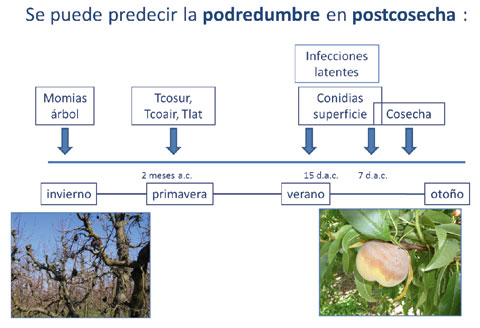
The incidence of post-harvest rot can predict from the moment in which appears the first latent infection and the conidia first of the pathogen on the surface of the fruit and in the air can be seen. The dates in which appear the first latent infection (Tlat), the first conidia of the pathogen on the surface of the fruit (Tcosur) and in the air (Tcoair) come to explain 99% of the incidence of the disease caused by ' Monilinia spp. ' on post-harvest.
The amount of conidia air present in the gardens and on the surface of the fruit varies in the Ebro Valley, according to the year of sampling, and also in terms of the phenological State of cultivation, being highest at the time of harvest. The density of the pathogen on the surface of the fruit during the last two weeks before harvest, explained 83 percent of these fruits in post-harvest rot.
Climatic conditions are critical to the infection of the fruits, temperature (T) and period of humidification (W), climatic factors that most influence on the penetration and infection of the fruits by ' Monilinia spp..' (Gell et al., 2008). In the case of immature fruit infection, which usually gives rise to infections that remain latent, these two climatic factors explain more than 90% of the variation in incidence (Gell et al., 2008), having more influence the duration of the period of moisture than temperature (Figure 8). In the Valley of the Ebro, the development of latent infections requires mild temperatures and moisture periods longer than 7 h (Figure 8), values that can be reached during the late spring and the end of the summer. In the case of rot of fruit at harvest, the incidence of the disease increases with temperature and the duration of the period of humidification. The optimum temperature for the infection of peaches by 'M. fructicola' during the harvest is from 22.5 to 25 ° C, with over 79% of the fruits are infected with periods of humidification of at least 12 h and temperatures below 27.5 ° C.
Figure 8: Surface obtained from the multiple regression model to predict the incidence of the latent infections in function of the temperature (T) and the period of humidification (W) (Gell et al., 2008).
Conclusions
The number of mummified fruit in the trees, latent infections, the density of inoculum of the pathogen in the air and on the surface of the fruit, the State fruit phenological, the values of temperature and moisture period, are critical to the choice of a strategy for control of the rot of the peach tree.
Bibliographic references
-Byrde, r. j., & Willetts, H. j. 1977. The Brown Rot Fungi of Fruit - Their Biology and Control. Pergamon Press, Oxford.
-Of lime, a. and Melgarejo, p. 2000. Mummified bone fruit (Monilinia spp..). In: Fruit nugget and bone diseases. E Montesinos, p. Melgarejo, M.A. Cambra and j. Pinochet (eds). Ed. Mundi-Prensa. Pp 66-67.
-EFSA Panel on Plant Health (PLH); Pest risk assessment of Monilinia fructicola for the US territory and identification and evaluation of risk management options. EFSA Journal 2011; 9 (4): 2119. [155 pp.] DOI:10.2903/j.EFSA.2011.2119. Available online: www.efsa.europa.eu/efsajournal
-Emery, k. M., Michailides, T. j. and Scherm, H. (2000). Incidence of latent infection of immature peach fruit by Monilinia fructicola and relationship to brown rot in Georgia. Plant Disease 84, 853-857.
-EPPO 2007. List of A2 pests regulated as quarantine pests in the EPPO region. OEPP/EPPO from http://www.eppo.org/QUARANTINE/listA2.htm.
-Gell, i., De Cal, a., Torres, r., Usall, j., & Melgarejo, p. 2008. Relationship between the incidence of latent infections caused by Monilinia spp.. and the incidence of brown rot of peach fruit: Factors affecting latent infection. European Journal of Plant Pathology 121: 487-498.
-Gell, i., De Cal, a., Torres, r., Usall, j., & Melgarejo, p. 2009. Conidial density of Monilinia spp.. on peach fruit surfaces in relation to the incidences of latent infections and brown rot. European Journal of Plant Pathology 123: 415-424.
-Landgraf, d. a., & Zehr, e. i. 1982. Inoculum sources for Monilinia fructicola in South Carolina peach orchards. Phytopathology 72: 185-190.
-Luo, y. and Michailides, T. j. 2001. Factors affecting latent infection of prune fruit by Monilinia fructicola. Phytopathology 91: 864-872.
-Ogawa, j. M., Zehr, e. i., and Biggs, a. r. 1995. Brown Rot. Pages 7-10 in: Compendium of Stone Fruit Disease. M. Ogawa, e. i. Zehr, g. w. Bird, D. f. Ritchie, k. Uriu, and j. k. Uyemoto, eds. American Phytopathological Society, St. Paul, MN.
-Villarino, M. Melgarejo, p., Usall, J. r., Segarra, j., and De Cal, a. 2010. Primary inoculum sources of Monilinia spp.. Spanish in peach orchards and their relative importance in Brown rot. Plant Disease 94: 1048-1054.
-Villarino, M., Sandin-Spain, p., Melgarejo, p., and De Cal, a. 2011. High Chlorogenic and Neochlorogenic Acid Levels in Immature Peach reduces Monilinia laxa Infection by come with Fungal Melanin Biosynthesis. Journal of Agricultural and Food Chemistry 59 (7), 3205-3213.
Responsable: Interempresas Media, S.L.U. Purpose: Subscription to our newsletter(s). User account management. Sending emails related to the same or related to similar or associated interests.Retention: for the duration of the relationship with you, or as long as necessary to carry out the specified purposesTransfer: Data may be transferred to other group companies for internal management purposes.Rights: Access, rectification, opposition, deletion, portability, limitation of processing and automated decisions: contact our DPD. If you consider that the processing does not comply with the regulations in force, you may lodge a complaint with the AEPD.More information: Data protection policy




")


 in tree 8 commercial orchards in the Valley of the Ebro...")
 on the floor of 8 gardens from the Ebro Valley...")