Mejora genética del espárrago
La base genética del espárrago cultivado es estrecha debido a que los actuales cultivares proceden de una misma población del S. XVIII (Purple Dutch). En su mayoría estos cultivares son diploides, verdes y de calibre grueso. Aunque es una especie dioica hoy día se tiende al desarrollo de variedades macho debido a las ventajas agronómicas que presentan las plantas macho. Desde 1999 se viene desarrollando en el Departamento de Genética de la Universidad de Córdoba un programa de mejora en espárrago con dos objetivos: por un lado desarrollar nuevas variedades poliploides con características diferentes (sabor, color, compuestos funcionales y morfología) y, por otro, se está generando un nuevo germoplasma empleando una raza local 4x (Morado de Huétor) y un conjunto de especies silvestres del género Asparagus. Es de esperar que este germoplasma pueda ser útil para ampliar la base genética del cultivo.
Introducción
El espárrago cultivado (Asparagus officinalis L.) es una especie monocotiledonea perteneciente a la Familia (Liliaceae) con un número de cromosomas 2n=20. Dentro del género Asparagus, A. officinalis es la única especie cultivada tanto para espárrago blanco como para espárrago verde. En la cuenca mediterránea se suelen recolectar los brotes jóvenes de poblaciones naturales de otras especies silvestres de este género (A. acutifolius L., A. aphyllus L., A. hórridus L., A. albus L.), comúnmente conocidos como espárragos trigueros y cuyo destino suele ser para autoconsumo o bien su comercialización en mercados locales.
La superficie cultivada de espárrago en el mundo es similar a la de otras hortícolas de importancia económica como el ajo, berenjena, judía y lechuga (FAOSTAT 2012). En España, Andalucía es la Comunidad Autónoma con mayor producción de esta hortaliza, siendo la Vega de Granada la principal zona productora de espárrago verde.

Colección de germoplasma del espárrago.
Historia del cultivo
Características botánicas

El sexo en espárrago está controlado por un solo gen (M/m). La dotación genética de las plantas macho es heterogamética Mm y la de las plantas hembra es homogamética mm (Löptien 1979). El sexo es una característica de interés en este cultivo puesto que las plantas macho son más productivas, precoces y longevas que las plantas hembra. Por el contrario, las plantas hembra dan lugar a espárragos o turiones de mayor calibre (Ellison, 1986). Por otro lado, el hecho de que las plantas macho no generen semilla supone una ventaja agronómica sobre las plantas hembra, puesto que las semillas actúan como mala hierba para el cultivo. Este mejor comportamiento agronómico de las plantas macho en cultivo ha provocado que la tendencia en la mejora de la especie vaya encaminada hacia el desarrollo de variedades macho.
Evolución varietal en espárrago
Se cree que fue a principios de la Edad Moderna cuando el espárrago alcanzó una gran difusión en los países de Europa Occidental. A principios del siglo XVIII, una población de origen holandés se fue haciendo cada vez más popular, la población conocida como Purple Dutch. A partir de esta población, mediante selección masal, se desarrollaron dos variedades base: Augenteüil, en Francia, y Gloria de Brunswick, en Alemania, que fueron posteriormente introducidas en Estados Unidos, derivándose de ellas distintos materiales en este país (Knaflewsky 1996). Casi todos los cultivares de espárrago utilizados en la actualidad proceden de estas variedades base. Sobre las variedades población base se ha llevado a cabo una larga labor de mejora desde principios del siglo XX, que conseguió incrementar la homogeneidad y el rendimiento de las distintas variedades población.
Antes de los años 70 no era posible explotar comercialmente las buenas combinaciones híbridas procedentes del cruzamiento entre una planta macho y otra hembra ya que una planta hembra genera como máximo unos pocos miles de semillas. Para poder ofrecer semilla en cantidad comercial se desarrollaron los llamados híbridos dobles que procedían del cruzamiento entre dos descendencias híbridas. Los híbridos dobles propiciaron un nuevo aumento del rendimiento y de la precocidad en comparación con las variedades población existentes (Corriols 1988). A partir de los setenta y con la puesta a punto del cultivo in vitro fue posible clonar las plantas parentales y se obtuvieron los llamados híbridos clonales simples o F1, procedentes del cruzamiento entre dos plantas clonadas para producir suficiente semilla. Esto permitió incrementar aún más la homogeneidad, precocidad y el rendimiento respecto a los híbridos dobles (Corriols 1988).
Debido a las ventajas que en cultivo presentan las plantas macho, hoy día la tendencia en el desarrollo de nuevas variedades va encaminada hacia la creación de híbridos clonales macho, compuestos sólo por plantas macho. Esto implica la obtención de plantas parentales llamadas Supermacho (MM) que se pueden obtener de dos formas: mediante la autofecundación de plantas macho andromonoicas o bien a través del cultivo in vitro de anteras. Los híbridos macho han tenido una importante difusión, inicialmente en la modalidad del cultivo para espárrago blanco y actualmente también en el cultivo para verde.
Objetivos en la Mejora Genética del espárrago
Entre las características de interés agronómico destacan el calibre de los turiones y la precocidad de la producción. En relación a las características morfológicas destacan dos objetivos en mejora genética: por un lado para su cultivo en verde interesa que la punta o yema terminal del espárrago permanezca cerrada hasta su recolección. Por otro lado, en el cultivo en blanco interesa que las variedades no generen compuestos antocianicos que dan lugar a coloraciones purpuras en las puntas de los turiones que empiezan a emerger por encima del caballón de cultivo.
La resistencia a estreses abióticos y bióticos es otro de los objetivos perseguidos en la mejora genética de esta especie. Entre los factores bióticos limitantes del cultivo destacan dos enfermedades: Fusariosis y Roya.
La fusariosis es la enfermedad más importante del cultivo para la que no existen tratamientos fitosanitarios de control eficaz. La fusariosis causa una podredumbre del rizoma y de las raíces y provoca el decaimiento prematuro de las plantaciones y además limita la replantación en suelos previamente infectados (Elmer, 2001). Las principales especies del genero Fusarium causantes de los problemas descritos son: Fusarium oxysporum f. sp. asparagi, F. moniliforme y F. roseum (Tello 1998).

La roya (Puccinia asparagi DC) es una enfermedad aérea que suele afectar a todas las zonas de producción, pero con una incidencia y severidad variable, dependiendo de las condiciones medioambientales. La búsqueda de resistencia a la roya fue el objetivo de uno de los programas de mejora llevados a cabo a principios del S.XX en EE UU, cuyo resultado dio lugar a la variedad Mary Washington que ha actuado de material de partida en todas las variedades mejoradas en EE UU, Canadá y en algunas desarrolladas en otras partes del mundo (Thiessen 1985, Chen et al. 1988).
Por otro lado, la incorporación de variabilidad genética en los programas de mejora es un objetivo también importante a tener en cuenta en la mejora genética de este cultivo. Se conocen algunas variedades locales tetraploides (2n=4x=40) cultivadas en España (Morado de Huétor), Italia (Violetto DAlbenga) y Argentina (Cereseto y Poire) que podrían ser empleadas para ampliar la base genética de los cultivares comerciales. Morado de Huétor es una variedad local cultivada en la Vega de Granada que presenta una alta variabilidad para caracteres agronómicos, morfológicos y organolépticos, y altos contenidos en compuestos funcionales (Moreno et al 2006, Fuentes-Alventosa et al 2008). Estas características pueden ser explicadas por el origen interespecífico (A. officinalis x A.maritimus) propuesto para esta raza local (Moreno et al., 2008). Morado de Huétor se caracteriza por dar lugar a espárragos con un calibre mediano-fino, un color variable que va desde el verde claro hasta el color morado y un sabor ligeramente amargo, a diferencia de los modernos cultivares que son muy homogéneos entre sí siendo estos en general de color verde, sabor dulce y calibre grueso.
Las especies próximas filogenéticamente pueden ser también un buen recurso genético. Se han obtenido híbridos entre la especie cultivada y algunas de estas especies: A. tenuifolius Lam. (Bozzini 1963), A. prostratus Dumort. (McCollum 1988) y A. maritimus Mill. (Falavigna et al. 2008) pero por el momento no se tiene constancia de su uso en mejora.
Mejora genética del espárrago en Córdoba (UCO IFAPA)
Desde 1999 se viene desarrollando en el grupo de Mejora Vegetal de la Universidad de Córdoba y en colaboración con el IFAPA (Alameda del Obispo -Córdoba) un programa de mejora genética en espárrago encaminado hacia la obtención de nuevas variedades poliploides y en el desarrollo de germoplasma que pueda ser útil para ampliar la base genética de las actuales variedades 2x (figuras 1 y 2). Para ello se está empleando la raza local Morado de Huétor, poblaciones naturalizadas de la especie cultivada y un conjunto de especies silvestres del género Asparagus spp. (Castro et al 2013).
Obtención de nuevas variedades

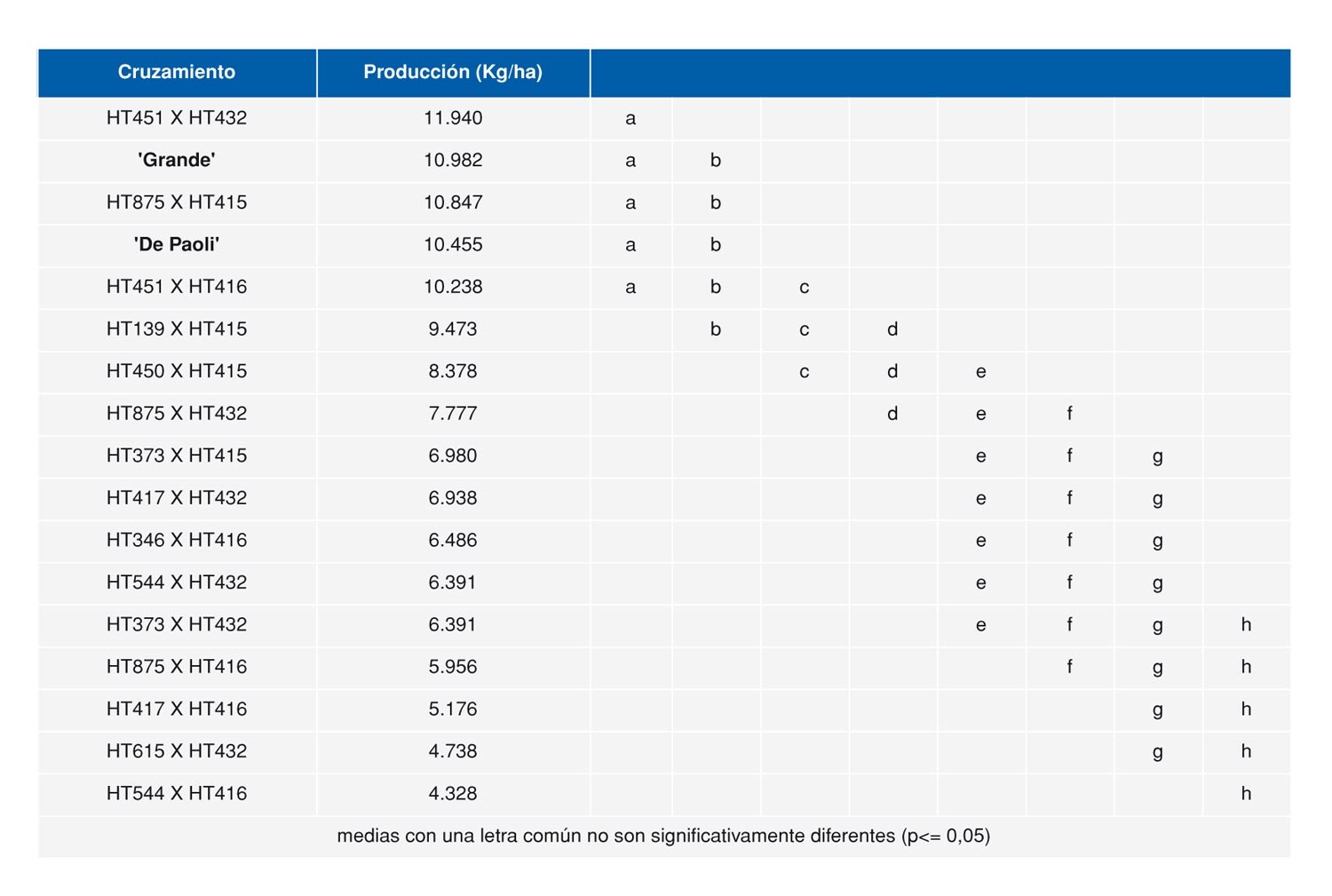
- Híbridos triploides: Las plantas seleccionadas como parentales por el Consejo Regulador y que posteriormente fueron caracterizadas como 3x mostraban además de un buen comportamiento agronómico un calibre superior al de la raza local y en el caso de las plantas hembra 3x se podía observar un alto grado de infertilidad. Estas observaciones sugirieron que podría ser interesante estudiar la posibilidad de obtener híbridos experimentales 3x en espárrago cruzando plantas 4x de la raza local con plantas 2x procedentes de cultivares comerciales. Un híbrido 3x obtenido de esta forma posibilitaría obtener híbridos con un comportamiento agronómico similar a los híbridos macho dada la infertilidad de las plantas y permitiría explorar además la posibilidad de explotar el vigor híbrido que en general se espera obtener al cruzar dos fondos genéticos diferentes. Hasta el momento se han llevado a cabo dos ensayos agronómicos con híbridos experimentales triploides que han puesto de manifiesto el elevado potencial productivo de estos híbridos (Moreno et al 2010; Moreno et al 2012b) (figura 3.). No obstante, no todas las buenas combinaciones híbridas detectadas pueden ser, por el momento, explotadas comercialmente debido a la baja fertilidad obtenida en algunos cruzamientos.
- Híbridos tetraploides: Los híbridos experimentales 3x evaluados en los ensayos mencionados anteriormente muestran un calibre intermedio entre la raza local 4x y los cultivares 2x. En las diferentes evaluaciones llevadas a cabo en los ensayos con híbridos 3x se observaron plantas que en muy baja frecuencia presentaban un calibre grueso similar a los cultivares comerciales. Las plantas hembra mostraban, además, un nivel de fructificación anormalmente alto para ser una planta triploide. El análisis ploídico de todas estas plantas reveló que eran tetraploides, en vez de triploides y el empleó de marcadores moleculares confirmó su condición híbrida. Por lo tanto estas plantas llevaban la mitad de su germoplasma de Morado de Huétor y la otra mitad de híbridos comerciales diploides. Algunas de estas plantas han sido empleadas en cruzamientos habiéndose observado un buen vigor híbrido. Las plantas macho tetraploides resultantes son en relación al sexo MMmm debido a que el fallo meiótico ocurrido en el parental 2x suele generar gametos MM ó mm (Camadro, 1994). Como consecuencia de esto generan una mayor proporción de plantas macho (83%) en el cruzamiento con plantas hembra (mmmm). Recientemente ha finalizado un ensayo agronómico con este tipo de híbridos experimentales 4x habiéndose obtenido en algunos de ellos un rendimiento y calibre similar a los testigos comerciales (tabla 1).
- Híbridos macho 4x: Durante el desarrollo del programa anteriormente mencionado se han realizado diferentes visitas a las parcelas plantadas con la variedad local Morado de Huétor encontrándose algunas plantas macho andromonoicas. Estos machos andromonoicos no mostraron características de interés agronómico y por ese motivo se cruzaron con hembras de élite seleccionadas en la raza local. Fruto de la selección de la descendencia se dispone de una colección de plantas macho andromonoicas que pueden ser un buen punto de partida para obtener plantas supermacho 4x (MMMm ó MMMM). Mediante la autofecundación de estas plantas y evaluación de sus descendencias se está intentando desarrollar una colección de plantas supermacho.

Mapa genético del espárrago
Paralelamente al trabajo anterior se están intentando aislar los 10 diferentes cromosomas del espárrago cultivado empleando la técnica citometría de flujo en colaboración con el Institute of Experimental Botany de Olomouc (República Checa). Aislar cromosomas individuales podría permitir asignar los grupos de ligamiento a cromosomas específicos y ser de ayuda en el establecimiento del cariotipo de la especie. Hasta el momento, se han obtenido 8 grupos de cromosomas que han podido ser aislados entre sí permitiendo el análisis individual de cada grupo.
Referencias bibliográficas
- Bozzini, A., 1963: Interspecific hybridization and experimental mutagenesis in breeding Asparagus. Genet Agrar 16, 212-218.
- Camadro EL, (1994). Second meiotic division restitution (SDR) 2n pollen formation indiploid and hexaploid species of Asparagus. Genet Resour Crop Ev 41: 1-7
- Castro, P., J. Gil, A. Cabrera, and R. Moreno, 2013: Assessment of genetic diversity and phylogenetic relationships in Asparagus species related to Asparagus officinalis. Genetic Resources and Crop Evolution 60, 1275-1288.
- Castro, P., J. Rubio, J. Gil, and R. Moreno, 2014: Introgression of new germplasm in current diploid cultivars of garden asparagus from a tetraploid spanish landrace Morado de Huétor. Scientia Horticulturae 168, 157-160.
- Chen, Y.W., Y.F. Yen and S.S. Chen, 1988: Breeding and varietal improvement developments and realases of Tainan(s) Nº1, Nº2, and Nº3. Proc. Fourth Symposium on Asparagus Res., Taiwan, 165-176.
- Corriols, L.,1988: Selección y mejora del espárrago. II Jornadas técnicas del espárrago, Pamplona, 27-37.
- Ellison, J.H., 1986: Asparagus breeding. Breeding Vegetable Crops, 521-569
- Elmer, W.H., 2001: Fusarium diseases of asparagus Fusarium: Paul E Nelson Memorial Symposium. APS Press, St-Paul, MN, 248-262.
- FAOSTAT-Agriculture 2012. Database. http://www.fao.org
- Falavigna, A., P. Alberti, P.E. Casali, L. Toppino, G. Mennella, and H. Wang, 2008: Interspecific hybridization for asparagus breeding in Italy Acta Horticulturae, 291-297, Vol. 776.
- Fuentes-Alventosa, J. M., Jaramillo, S., Rodríguez-Gutiérrez, G., Cermeño, P., Espejo, J. A., Jiménez-Araujo, A., ... & Rodríguez-Arcos, R. (2008). Flavonoid profile of green asparagus genotypes. Journal of agricultural and food chemistry, 56(16), 6977-6984.
- Gonzalez, A., J.A. Fernandez, and S. Bañón, 1993: Cultivo de espárrago verde en invernadero., In: Mundi-Prensa., (ed.), 12-36.
- Johnson, D.A., and J. Lunden, 1992: Effect of rust on yield of susceptible and resistant asparagus cultivars. Plant disease 76, 84-86.
- Knaflewski, M., 1996: Genealogy of asparagus cultivars, 87-91, Vol. 415.
- Löptien, H., 1979: Identification of the sex chromosome pair in asparagus (Asparagus officinalis L.). Zeitschrift für Pflanzenzüchtung.
- Luný, J., 1979: The history of asparagus as a vegetable, the tradition of its growing in Czechoslovakia (CSSR) and the prospect of its further propagation and breeding. Proceeding of the 5th International Asparagus Symposium, G. Reuther. Geisenheim. In: Eucarpia (ed.), 82-86.
- McCollum, G., 1988: Asp 8271 and Asp 8284 asparagus germplasm. HortScience (USA).
- Moreno, R., J.A. Espejo, A. Cabrera, T. Millán, and J. Gil, 2006: Ploidic and molecular analysis of 'Morado de Huetor' asparagus (Asparagus officinalis L.) population; a Spanish tetraploid landrace. Genetic Resources and Crop Evolution 53, 729-736.
- Moreno, R., J.A. Espejo, A. Cabrera, and J. Gil, 2008: Origin of tetraploid cultivated asparagus landraces inferred from nuclear ribosomal DNA internal transcribed spacers' polymorphisms. Annals of Applied Biology 153, 233-241.
- Moreno, R., J.A. Espejo, and J. Gil, 2010a: Development of triploid hybrids in asparagus breeding employing a tetraploid landrace. Euphytica 173, 369-375.
- Moreno R., Castro P., Rubio J., Rodríguez-Arcos R., Gil J. 2012a. Desarrollo de una nueva variedad de espárrago octoploide HT801. Actas de Horticultura 60: 105-108.
- Moreno R., Rubio J., Castro P., Gil J. 2012b. Evaluación del rendimiento de híbridos experimentales triploides en espárrago. Actas de Horticultura 62: 147-148
- Moreno R., García T., Cabrera A., Millán T., Gil J., Vrána J., Kubaláková M., Číhalíková, J., Doleel J. (2014). Contribución al mapa genético del espárrago (Asparagus officinalis L.). Actas de Horticultura 69: 181-182.
- Rigau, A., 1988: Cultivo de espárragos, 35-44 Ed. Sintes.
- Tello, J.C., 1988: Las fusariosis del espárrago. II Jornadas Técnicas del espárrago, Pamplona, 63-77.
- Thiessen, H., 1985: Cultivar evaluation in Canada. Proceedings of the VIth International Asparagus Symposium, Guelph, 63-72.
